Депонировано в ВИНИТИ, рег. №. 1607-В89 от
16.02.1989, 19 с.
Жизнь млекопитающих: энергетический анализ
Федосин С.Г.
Энергетический анализ используется для нахождения формул, описывающих
метаболизм млекопитающих, их пищевой рацион, время жизни, воспроизведение
потомства и другие основные жизненные характеристики.
В понятие метаболизма мы включаем обмен веществ и рассматриваем его в
терминах затраты энергии и пищи, необходимых животным для их существования.
Энергия, получаемая в среднем за сутки, затрачивается животным на перемещение в
пространстве, поддержание и функционирование органов и тканей, а также
(у млекопитающих и птиц) – на поддержание постоянной температуры тела. Баланс
мощности как баланс энергии за сутки, имеет вид:
![]() (1)
(1)
где ![]() – общая мощность,
– общая мощность,
![]() – мощность,
затрачиваемая для перемещения в пространстве,
– мощность,
затрачиваемая для перемещения в пространстве,
![]() – мощность питания
органов и тканей тела, включая работу
дыхания и сердца,
– мощность питания
органов и тканей тела, включая работу
дыхания и сердца,
![]() – мощность
теплопотерь,
– мощность
теплопотерь,
![]() – коэффициент,
показывающий степень превращения части химической энергии пищи в энергию,
используемую организмом,
– коэффициент,
показывающий степень превращения части химической энергии пищи в энергию,
используемую организмом,
![]() – масса пищи в
килограммах.
– масса пищи в
килограммах.
Выразим все компоненты общей мощности в (1) как степенные функции от
массы тела животного ![]() , учитывая, что скорость передвижения животных
пропорциональна массе тела в степени
0,17 , а теплопотери пропорциональны массе тела в степени 0,5:
, учитывая, что скорость передвижения животных
пропорциональна массе тела в степени
0,17 , а теплопотери пропорциональны массе тела в степени 0,5:
![]()
![]()
![]()
![]() (2)
(2)
где ![]() – некоторые константы,
которые мы далее определим.
– некоторые константы,
которые мы далее определим.
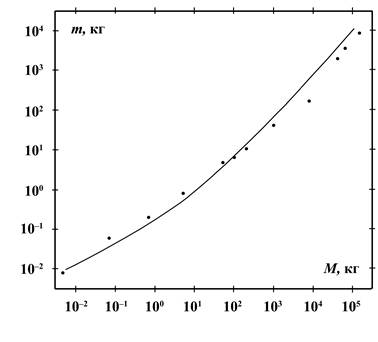
На рисунках 1 и 2 приведены зависимости массы съедаемой пищи в сутки,
усреднённые за год, от массы тела у 48 видов плотоядных и 24 видов
растительноядных животных. Кривые проведены таким образом, чтобы они
удовлетворяли соотношению (2) и известным из наблюдений суточным рационам, в
случае плотоядных животных самым маленьким видом были землеройки, а самым
большим – киты. Это позволило прокалибровать соотношение (2) и найти
коэффициенты ![]() для плотоядных
животных следующим образом:
для плотоядных
животных следующим образом:
![]() (3)
(3)
Из (3) следует, что плотоядное животное при массе ![]() кг имеет
среднесуточный рацион в размере
кг имеет
среднесуточный рацион в размере ![]() кг.
кг.
Зависимость рациона для растительноядных животных имеет тот же вид, что
и (3), за исключением того, что вместо коэффициента 0,011 стоит коэффициент
0,016.
Это означает, что при той же массе растительноядное животное поглощает пищи
в 0,016/0,011 = 1,45 раз больше, что
является следствием меньшей питательности растительной пищи по сравнению с пищей
животной.
Исследования показывают, что в экосистемах виды обычно располагаются
так, что каждый из них получает относительно одинаковые пищевые ресурсы.
Математически это означает, что плотность популяции того или иного вида зависит
обратно пропорционально от суточного рациона. Обозначим через ![]() число особей
популяции, а через
число особей
популяции, а через ![]() – площадь территории
экосистемы, тогда имеем:
– площадь территории
экосистемы, тогда имеем: ![]() ~
~ ![]() или
или ![]() ~
~ ![]() . В последнем равенстве справа стоит неизменная площадь экосистемы,
а слева – общий суточный рацион той популяции, чьи особи имеют суточный
рацион
. В последнем равенстве справа стоит неизменная площадь экосистемы,
а слева – общий суточный рацион той популяции, чьи особи имеют суточный
рацион ![]() . Следовательно, популяция каждого вида имеет один и тот же общий
суточный рацион, независимо от величины массы тела животных, и приблизительно в
одинаковой степени эксплуатирует биологические ресурсы экосистемы.
. Следовательно, популяция каждого вида имеет один и тот же общий
суточный рацион, независимо от величины массы тела животных, и приблизительно в
одинаковой степени эксплуатирует биологические ресурсы экосистемы.
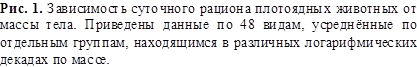
Соотношение (3) можно применить для калибровки так называемой кормовой
единицы, определяющей энергетическую ценность корма сельскохозяйственных
животных. Оказывается, что средний суточный рацион в кормовых единицах имеет
следующую зависимость от массы животных:
![]()
Здесь коэффициент 0,2 отражает тот факт, что данные животные находятся
в закрытых помещениях и имеют в среднем пониженные в 5 раз энергетические
траты, связанные с движением. Сравнение с (3) позволяет заключить, что одна
кормовая единица эквивалентна 0,011/0,004 = 2,75 кг животной пищи и 0,016/0,004
= 4 кг растительной пищи.
![]()

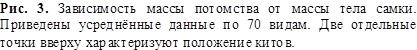
Уравнение энергетического баланса (3) позволяет понять многие факты из
жизни млекопитающих. Например, рисунок 3 показывает зависимость массы потомства
![]() , вычисляемой как произведение массы одного детёныша на
среднее количество детёнышей, от массы самки, по 70 видам животных. Проведённая
кривая хорошо аппроксимирует данные в огромном диапазоне масс и описывается
уравнением, подобным (3):
, вычисляемой как произведение массы одного детёныша на
среднее количество детёнышей, от массы самки, по 70 видам животных. Проведённая
кривая хорошо аппроксимирует данные в огромном диапазоне масс и описывается
уравнением, подобным (3):
![]() (4)
(4)
Отсюда следует, что масса потомства пропорциональна количеству энергии,
поступающей в организм матери с пищей, и составляет от неё всегда одну и ту же
долю (в условных единицах), независимо от массы животных.
Другой пример касается интенсивности метаболизма, пропорциональной
потреблению кислорода животным в единицу времени. Кислород используется в
организме в конечном счёте для окисления составных частей пищи и получения тем
самым химической энергии.
Обычно показатель интенсивности метаболизма вычисляется на основании
данных, полученных при исследовании голодных животных в состоянии покоя –
измеренное количество потреблённого кислорода переводится в соответствующее
количество получающейся химической энергии за единицу времени из расчёта, что 1
литр кислорода соответствует 20,1 кДж. График зависимости интенсивности
метаболизма от массы тела у различных млекопитающих и птиц, построенный в
логарифмическом масштабе, представляет собой почти прямую линию. Эта загадочная
зависимость носит название «кривая от мыши до слона» [1] и аппроксимируется
следующим уравнением:
![]() (5)
(5)
Многие исследователи указывают на то, что (5) не имеет ясного смысла –
непонятно, почему показатель степени должен равняться 0,74. Поскольку интенсивность
метаболизма ![]() измеряется в ваттах и
фактически есть энергетическая мощность, мы можем использовать вместо (5)
уравнение баланса (2) с коэффициентами из (3). Учитывая ещё, что для
неподвижных животных энергия передвижения
измеряется в ваттах и
фактически есть энергетическая мощность, мы можем использовать вместо (5)
уравнение баланса (2) с коэффициентами из (3). Учитывая ещё, что для
неподвижных животных энергия передвижения ![]() равна нулю, для
интенсивности метаболизма должно быть:
равна нулю, для
интенсивности метаболизма должно быть: ![]() , где
, где ![]() – некоторый
коэффициент.
– некоторый
коэффициент.
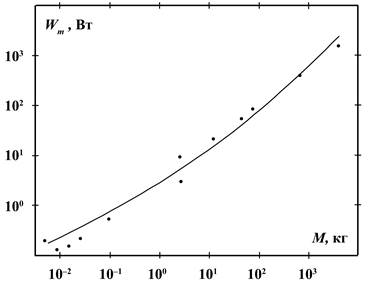

На рисунке 4 приведены данные по интенсивности метаболизма в виде точек
на графике, которые можно описать следующим соотношением:
![]() (6)
(6)
Соотношения (5) и (6)
имеют приблизительно одинаковую точность в середине диапазона масс, однако (6)
лучше на краях диапазона, где у (5) наблюдаются отклонения – увеличенные в 3
раза у землероек и в 2 раза у китов интенсивности метаболизма. Например, для Sorex cinereus
при её массе 0,003 кг интенсивность метаболизма согласно (5) должна быть 0,048
Вт, из (6) получается 0,13 Вт, а опытное значение равно 0,15 Вт [2]. Согласно
(6) интенсивность метаболизма покоящихся животных определяется потерями энергии
на питание тела и на его обогрев.
Количество потреблённого кислорода и интенсивность метаболизма
покоящихся животных зависит как от массы тела, так и от частоты дыхания – чем
больше масса и частота дыхания, тем больше будет величина ![]() . Следовательно, для частоты дыхания в минуту можно записать:
. Следовательно, для частоты дыхания в минуту можно записать:
![]() ~
~ ![]() или
или
![]() (7)
(7)
Полученное соотношение более обоснованно, чем часто используемая
биологами [3] простая степенная зависимость вида ![]() , и точнее на краях диапазона масс. Аналогично для частоты
сокращения сердца в минуту у находящихся в состоянии покоя млекопитающих вместо
степенной зависимости вида
, и точнее на краях диапазона масс. Аналогично для частоты
сокращения сердца в минуту у находящихся в состоянии покоя млекопитающих вместо
степенной зависимости вида ![]() должно быть:
должно быть:
![]() (8)
(8)
Коэффициенты 2,07 в (7) и 9,3 в (8) были выбраны из условия наилучшего
соответствия имеющимся данным. Надо заметить, что вблизи самых малых масс
точность (8) уменьшается, поскольку у землероек и мышей вместе с частотой
биения сердца ещё более значительно увеличиваются относительные размеры сердца,
что в сумме и обеспечивает их метаболизм.
Обратимся теперь к временным характеристикам – продолжительности жизни ![]() , возрасту полового созревания
, возрасту полового созревания ![]() и продолжительности
беременности
и продолжительности
беременности ![]() млекопитающих. Все эти
характеристики явно коррелируют между собой. С другой стороны,
продолжительность жизни явно зависит от двух факторов: она больше при
пониженном энергетическом обмене (что характерно для долгожителей) и
увеличивается с ростом массы животного (чем больше масса, тем дольше надо её
наращивать). Математически это можно записать в виде пропорциональной
зависимости:
млекопитающих. Все эти
характеристики явно коррелируют между собой. С другой стороны,
продолжительность жизни явно зависит от двух факторов: она больше при
пониженном энергетическом обмене (что характерно для долгожителей) и
увеличивается с ростом массы животного (чем больше масса, тем дольше надо её
наращивать). Математически это можно записать в виде пропорциональной
зависимости: ![]() , где
, где ![]() – масса тела
животного,
– масса тела
животного, ![]() – мощность обмена согласно (2). Данная зависимость
превращается в равенство с учётом коэффициентов из (3) и данных из рисунка 5,
где представлена продолжительность жизни (в годах) млекопитающих в зависимости
от массы тела:
– мощность обмена согласно (2). Данная зависимость
превращается в равенство с учётом коэффициентов из (3) и данных из рисунка 5,
где представлена продолжительность жизни (в годах) млекопитающих в зависимости
от массы тела:
![]() (9)
(9)
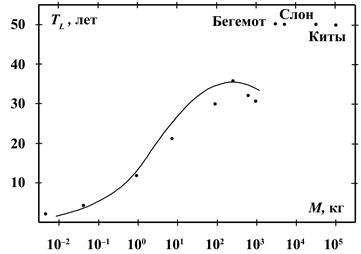
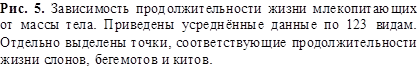
Из (9) следует, что при больших массах (более 200 кг) продолжительность
жизни начинает уменьшаться, поскольку возрастают потери энергии на передвижение
животного. Если же пищи достаточно и не надо затрачивать много энергии на её
поиск, то такие животные живут больше обычного – таковы приматы, слоны, бегемоты
и носороги, живущие в тёплых странах. Летучие мыши 90 % своей жизни неподвижны
и экономят на энергии движения, охотясь лишь в сумерки вечером и утром. Они
имеют увеличенную продолжительность жизни ещё и потому, что минимизируют свои
теплопотери зимой во время спячки. Краткий список типичных долгожителей
некоторых видов живых существ приведён в Таблице 1.
Таблица 1
Продолжительность жизни некоторых видов – долгожителей.
|
Название |
Продолжительность жизни, годы |
|
Губка |
50 |
|
Актиния |
85 |
|
Жемчужница |
100 |
|
Речной
рак |
30 |
|
Сетчатый
питон |
70 |
|
Слоновая
черепаха |
175 |
|
Сосна |
400 |
|
Арча |
2000 |
|
Попугай
какаду |
70 |
|
Бычий
солитёр |
18 |
|
Серая
жаба |
40 |
|
Летучая
мышь |
24 |
Если не учитывать такие факторы, как наличие врагов, достаточность пищи
или склонность к заболеваниям, главной особенностью, объединяющей всех
долгожителей, является пониженный энергетический обмен. Арча, растущая на
горных склонах, обязана своим долголетием бедному составу питательных веществ,
находящихся в каменистых почвах. Привольно живётся и паразитам, нашедшим
надёжного хозяина, и имеющих рядом и стол и дом.
Продолжительность жизни китообразных в принципе также может быть
описана с помощью выражения типа (9), но только с другими коэффициентами,
поскольку у водоплавающих млекопитающих другие зависимости отдельных компонент
мощности в соотношениях (1) и (2). В поисках пищи эти животные передвигаются в
воде не только горизонтально, но в не меньшей степени и вертикально, совершая
работу против небольшой по величине силы сопротивления воды и значительной
выталкивающей силы Архимеда. Работа против силы Архимеда может быть вычислена
как работа силы тяжести, она пропорциональна массе тела. Если в знаменателе
соотношения (9) член ![]() заменить на
заменить на ![]() и устремить массу
и устремить массу ![]() к бесконечности, то
для такой величины продолжительности жизни самых массивных морских животных
получим кривую зависимости с насыщением:
к бесконечности, то
для такой величины продолжительности жизни самых массивных морских животных
получим кривую зависимости с насыщением:
![]() лет,
лет,
что вполне соответствует положению дел с возрастом китов.
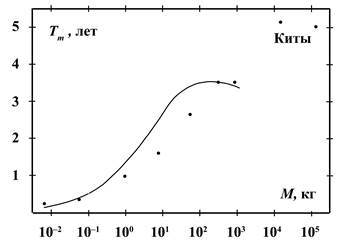
На рисунках 6 и 7 приведены усреднённые данные о возрасте полового
созревания самок (в годах) и продолжительности их беременности (в месяцах) в
зависимости от массы тела. Эти данные описываются соотношениями, подобными (9):
![]() (10)
(10)
![]() (11)
(11)
![]()
Из (9) и (10) следует, что продолжительность жизни приблизительно в 10
раз превышает возраст полового созревания. Поскольку у человека в 15 лет
практически заканчивается развитие основных половых признаков, то отсюда
следует, что продолжительность жизни человека должна быть 150 лет.
В интервале масс от 1 до 200 кг данные о продолжительности беременности
на рисунке 7 находятся несколько ниже, а
при массах от 200 до 1000 кг – выше, чем кривая по соотношению (11). Это можно
объяснить приспособлением к смене времён года. Например, зайцы в течение
благоприятного сезона могут иметь 2 – 3 выводка. Малая продолжительность
беременности в данном случае помогает переносить голодную и трудную зиму.
Животные с массой более 200 кг обычно имеют беременность более 7 месяцев и
потому захватывают зимний период. Поэтому увеличение срока беременности
позволяет иметь более здоровое и подготовленное к жизни потомство, особенно у
копытных, не имеющих убежищ.
Приведённые выше результаты тесно связывают между
собой потоки носителей, энергии и упорядоченности. На примере млекопитающих
видно, как поток носителей в виде массы пищи преобразуется в живых существах в
поток энергии, позволяющий им перемещаться в пространстве, упорядочивать свою
внутреннюю структуру и окружение, сохранять свою организацию с помощью
гомеостаза. Развитый энергетический подход легко может быть применён ко всем видам
живого с учётом их индивидуальных особенностей. Например, интенсивность метаболизма и скорость
продукции фотосинтеза у сосудистых растений изменяются по тому же закону, что и
интенсивность метаболизма у животных, с аналогичной зависимостью от массы тела,
одинаковы и зависимости плотности популяции [4].
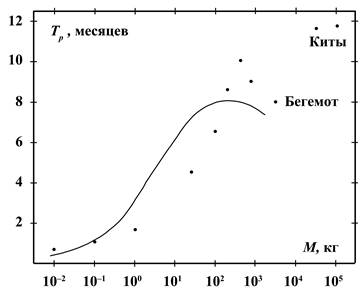
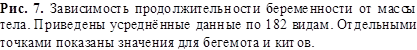
На рисунках 5, 6, 7 имеются экстремальные значения
продолжительности жизни, возраста полового созревания и продолжительности
беременности. Они могут быть объяснены разной структурой распределения
энергетических потоков, зависящей от массы тела, количества клеток в организме
и количества связей между ними, от устройства внутренней организации, от
соотношения между объёмом и поверхностью тела (что важно для оценки теплопотерь
при поддержании постоянной температуры тела). Следовательно, при прочих равных
условиях некоторые виды (в том числе человек) получают преимущество в своём
развитии – их экологическая ниша и внутренняя организация оказываются такими,
что при относительно небольших затратах энергии и требуемых запасах питания
становится возможным иметь наибольшую продолжительность жизни. Последнее же
является основным для приобретения жизненного опыта и передачи его потомкам – в
виде упорядочения среды своего обитания (искусственная среда, техносфера) и в
виде жизненно важной информации.
Из соотношений (2), (3) и (6) следует следующая
зависимость между интенсивностью метаболизма и массой тела свободно живущих
сухопутных млекопитающих:
![]() (12)
(12)
Умножая интенсивность метаболизма (12) на
продолжительность жизни согласно (9) и переводя годы жизни в секунды, приблизительно
оценим энергию в джоулях, которую использует организм в течение всей своей
жизни:
![]() или
или
![]()
То, что энергия, проходящая через организм и
используемая им, пропорциональна массе тела ![]() , означает следующее: каждая единица массы любого животного
пропускает через себя одно и то же количество энергии в течение своей жизни,
удельная энергия равна константе. Однако, как было сказано выше, эффективность
использования этой энергии совсем не одинакова – у доминирующих видов энергия
используется с большей выгодой для них.
, означает следующее: каждая единица массы любого животного
пропускает через себя одно и то же количество энергии в течение своей жизни,
удельная энергия равна константе. Однако, как было сказано выше, эффективность
использования этой энергии совсем не одинакова – у доминирующих видов энергия
используется с большей выгодой для них.
С помощью (12) можно оценить процентную долю каждой из
компонент энергии для среднего человека с
массой M = 65 кг в общем балансе его
энергии. Первая компонента энергии в (12), описывающая затраты на движение,
даёт 32 %, вторая компонента связана с питанием тела и вносит вклад 45 %, тогда
как последняя компонента, ответственная за обогрев тела, равна 23 %. Интересно
сравнить эти цифры с затратами энергии различного вида в масштабе целой страны.
Например, в [5] приведены данные о потреблении энергии в США в 1995 году. Затраты
энергии на транспорт составили около 27 %; далее можно выделить затраты
промышленности 38 %, продажи пищи 2,6 %
и её домашнего
приготовления 5 % (всё вместе около 46 % как аналог для потребления
общества); наконец, затраты энергии на жилищное хозяйство и коммунальные услуги
– до 27 %. Сравнивая затраты для одного
человека и затраты целой страны, находим их близкое соответствие, что можно
трактовать как повторение структуры энергетических потоков отдельного человека
на более высоком – общественном уровне.
Оценим, сколько характерных событий типа дыхательных циклов и ударов
сердца происходит в течение жизни млекопитающего. Если в метаболизме животных
учесть движение, то в среднем частота дыхания в минуту увеличится. Заменяя в
(7) интенсивность метаболизма покоя ![]() на энергетическую
мощность
на энергетическую
мощность ![]() из (12), найдём
усреднённую частоту дыхания свободно живущего животного, а умножая её на время
жизни (9) в минутах, определим среднее количество дыхательных циклов за всю
жизнь:
из (12), найдём
усреднённую частоту дыхания свободно живущего животного, а умножая её на время
жизни (9) в минутах, определим среднее количество дыхательных циклов за всю
жизнь:
![]() ,
,
![]()
Аналогично, для частоты усреднённого сердечного ритма в минуту у
свободно живущего животного вместо (8) имеем:
![]()
Если умножить эту частоту сердца на время жизни (9) и на 5,26ּ105 минут, содержащихся в каждом годе жизни,
можно подсчитать общее количество ударов сердца:
![]()
Средние количества таких событий, как дыхательные и сердечные циклы,
оказываются одинаковыми для всех млекопитающих. Биологические часы животных,
связанные с их внутренними циклами, с их физиологией и метаболизмом, хотя и
идут с разными скоростями, но имеют один и тот же предел своих показаний.
Ответим теперь на вопрос, как с молекулярной точки
зрения соотносится количество пищи, необходимое организму, с временными
циклами. Поскольку основной вклад в молекулярную массу млекопитающих вносит
вода (в организме человека 65 %
всей массы даёт кислород, 18 % – углерод и 10 % – водород) как одна из самых
простых молекул, то в первом приближении общая тепловая энергия всего тела
определяется именно водой. Температуры тела млекопитающих лежат в основном в
интервале значений от 36 до 40 °С, причём плацентарные млекопитающие
характеризуются температурой 38 °С, сумчатые – 36 °С, однопроходные типа ехидны
и утконоса – 30 °С. У птиц температура тела несколько выше – около 40 °С. В
градусах Кельвина средняя температура млекопитающих будет равна ![]() К. При такой
температуре молекулы воды имеют скорость
К. При такой
температуре молекулы воды имеют скорость ![]() и среднюю кинетическую
энергию движения в виде:
и среднюю кинетическую
энергию движения в виде: ![]() , где
, где ![]() – масса молекулы воды,
– масса молекулы воды,
![]() – постоянная Больцмана.
Следовательно, тепловая энергия (в джоулях), заключающаяся в организме с массой
– постоянная Больцмана.
Следовательно, тепловая энергия (в джоулях), заключающаяся в организме с массой
![]() , исчисляемой в килограммах, будет равна:
, исчисляемой в килограммах, будет равна:
![]() или
или ![]() (13)
(13)
Из выражения (12) также можно найти отношение энергии
к массе тела. Поскольку выявлено, что базисный ритм временной организации живых
систем колеблется вблизи суток, возьмём в качестве характерного цикла обменных
процессов у млекопитающих суточный период, равный величине ![]() секунд. Тогда
произведение интенсивности метаболизма
секунд. Тогда
произведение интенсивности метаболизма ![]() из (12) на
из (12) на ![]() даст энергию, которой
обладает животное в течение суток. Отношение этой энергии к массе тела будет равно:
даст энергию, которой
обладает животное в течение суток. Отношение этой энергии к массе тела будет равно:
![]() (14)
(14)
Величина ![]() равна: при массе тела
10–3 кг – 3,8·106, при массе 1 кг – 2,7·105,
при массе 103 кг – 1,1·105
равна: при массе тела
10–3 кг – 3,8·106, при массе 1 кг – 2,7·105,
при массе 103 кг – 1,1·105 ![]() . Если сравнить эти данные с (13), то видно, что маленькие
млекопитающие за сутки получают с пищей гораздо больше энергии, чем их
собственная тепловая энергия. Для животных с массой порядка 1 кг суточный поток
энергии сравним по величине с теплосодержанием в организме, а крупным
млекопитающим для приобретения того уровня энергии, который позволил бы им
восстановить необходимый уровень организации в случае необходимости, суток уже
не хватает, и их обменный цикл длиннее. Однако если перевести всех животных в
неподвижный режим и в лабораторную обстановку, в которой нет теплопотерь, то
первый и третий члены в скобках соотношения
(14) исчезают, и мы получим:
. Если сравнить эти данные с (13), то видно, что маленькие
млекопитающие за сутки получают с пищей гораздо больше энергии, чем их
собственная тепловая энергия. Для животных с массой порядка 1 кг суточный поток
энергии сравним по величине с теплосодержанием в организме, а крупным
млекопитающим для приобретения того уровня энергии, который позволил бы им
восстановить необходимый уровень организации в случае необходимости, суток уже
не хватает, и их обменный цикл длиннее. Однако если перевести всех животных в
неподвижный режим и в лабораторную обстановку, в которой нет теплопотерь, то
первый и третий члены в скобках соотношения
(14) исчезают, и мы получим:
![]() (15)
(15)
Данная величина в 4 раза меньше, чем (13). Это даёт
основания полагать, что любому животному необходимо около 4 суток для
накопления энергии с целью молекулярного переустройства внутренней организации
в тканях и органах. Из (15) следует, что каждый килограмм тела покоящегося
животного требует для своего питания энергии не менее 4,9·104 Дж в
сутки. Поскольку оценка массы всех животных Земли даёт величину 2·1012
кг, то в год все они потребляют порядка 3,5·1019 Дж энергии. Как и
следовало ожидать, это количество энергии меньше глобальной мощности
фотосинтеза, достигающей по данным из [6] значения 9·1021 Дж в год.
Литература
1. Шмидт-Ниельсен К. Размеры животных: почему они
так важны? – М., Мир, 1987.
2.
Morrison P.R., Ryser F.A., Dawe A.R. Studies on the
physiology of the masked shrew Sorex cinereus. – Physiol. Zool., 1959, V. 32,
P. 256 – 271.
3.
Stahl W.R. Scaling
of respiratory variables
in mammals. – J. Appl. Physiol.,
1967, V. 22, P. 453 – 460.
4.
Enquist B.J., Brown J.H., West G.B. Allometric scaling
of plant energetics and population density. – Nature, 1998, V. 395, 10 September,
P. 163 – 165.
5. Путвинский С.В. Возможна ли будущая мировая энергетическая
система без ядерного синтеза? – Успехи
физических наук, 1998, Т. 168, № 11, С. 1235 – 1246.
6. Изаков М.Н. Самоорганизация и информация на планетах и
в экосистемах. – Успехи физических наук,
1997, Т. 167, № 10, С. 1087 – 1094.
7. Шмидт-Ниельсен К. Физиология животных: приспособление
и среда. – М., Мир, 1982.
Источник:
http://sergf.ru/zh.htm